|
|
|
|
|
|
|
|
||
|
|
|||
|
|
|||
|
|
|||
|
|
|||
|
|
|||
|
REVIEW DRAFT 3/10/03 Transmissible Postulates, Copyright March, 2003 Chronic Wasting Disease: Observations on TSE Transmissibility R.A. Forrest The CWD Foundation, Box 55, South Fork, Colorado 81152 Article best viewed using Acrobat Reader. Abstract: (Part IIc) Transmissible Spongiform Encephalopathies (TSE) and in particular, Chronic Wasting Disease (CWD) are devastating neuropathologic diseases caused by a unique, but unknown infective agent with high degree of disinfectant refractivity. Collective literature review and animal observational data has defined two prospective varieties or stains of the infective CWD agent, both suspected to be caused by Spiroplasma bacteria. Strain "CWDe" is ensconced in an insect vector, most likely astigmatid grain, forage, or hay mites common in pastures or hay fields. CWDe affects both young and old cervids and is fully dosage dependant, environmentally resilient, and capable of sustaining cervid infections over many years. CWDe has few symptoms apparent prior to clinical disease. "CWDr" is more of a classic respiratory infection mechanism either as a direct Spiroplasma bacterial infection or S. spp. attendant with a mycoplasma commensal agent. Symptoms are pronounced in young animals resulting in severe neonate mortality and runting syndrome prior to demonstrating terminal CWD symptoms many months later. Astigmatid mites give the CWDe strain a second plausible infection route via ecto-parasitic mites of the genus Psoroptes. P. ovis and P. cuniculi are believed to be the same species and P. cuniculi has been found at all heavily infected CWD facilities. As Spiroplasma is believed to be present in lymph, the Psoroptes spp. can collect CWD infectivity, harbor and reproduce infectivity, and then deliver it to a new host, thereby spreading disease. The consistent presence of Psoroptes infestations coincident with CWD infections must place Psoroptes as a serious potential TSE transmission vector. INTRODUCTION Chronic Wasting Disease (CWD) is a Transmissible Spongiform Encephalopathy (TSE) affecting both wild and domestic deer species. The causative agent is unknown, but possesses very unconventional biological and physical properties. A more conventional biological agent, such as the enigmatic Spiroplasma bacteria has been offered (Forrest, 2002, Part I). Unusually long incubation periods, progressive central nervous system degeneration and the characteristic histopathological lesions distinguish TSEs. Only minimal or insignificant immune or inflammatory response is generally present. While considerable controversy concerns the nature of the TSE causative agent, spontaneous, genetic and transmissible varieties have been described. In Forrest (2002) Part I, "A Logical Causative Agent" evidence demonstrated the fallibility of current testing procedures and that Spiroplasma, a mycoplasma derived from the Class Mollicutes, retains the distinction of harmonizing with the most known TSE agent characteristics. In Forrest (2003a) Part IIa, "Novel Vectors" the relationship of insects, Spiroplasma and TSE was examined. In Part IIb "The Chronic Conundrum" (Forrest, 2003b) wild and domestic case studies linked puzzling and seemingly random or co-incidental events into a consistent, cohesive picture of disease transmission characteristics. The fully defined pattern is conducive to a Spiroplasma infection. Spiroplasmas, like all mycoplasmas are parasitic, difficult-to-culture bacteria lacking in cell walls. They have an affinity for mammalian membranes, and an appetite for sterol and phospholipids. Spiroplasma bacteria are capable of producing highly oxidant hydrogen peroxide byproducts, and will seemingly metabolize parts of the host's immune system. Importantly, they possess a shape-shifting character entering an intracellular "stealth" mode making detection by microscopy difficult. Astigmatic grain or storage mites have been implicated in the potential transmission of TSE. Spiroplasmas are likely to be present in infective mites. Cervids grazing upon infected fields, or ingesting mite-infected hay may obtain sufficient pathogen to result in a TSE infection (Forrest, 2003b). Currently, one of the preferred methods of experimental TSE infection is via ingestion of diseased tissue. As documented by Sigurdson (1999), experimental CWD infections have shown symptomatic abnormal prion protein (PrPres) development within alimentary-tract-associated lymph nodes as early as 42 days after oral inoculation. The gastrointestinal tract essentially appears to be one natural route of TSE infection. The mode of TSE transmission is presently unknown. One must note that the following precepts as presented herein are hypothetical, yet are founded upon documented and observable phenomena. The mechanisms and concepts herein are predicated upon solid literature review, conceptual probability, likeness to other more conventional disease mechanics, and observational data evaluation. Significant supplementary field investigative work with its attendant laboratory effort must be completed prior to firmly establishing a true causal agent-transmission linkage. Please entertain the following presentation as possible working hypotheses as to the nature, resilience and consequential actions of the Chronic Wasting Disease causative agent and its environmental vectors. POSTULATED CWD STRAINS The extremely anomalous and co-incidental presence of high neonatal cervidea mortality at highly CWD-contaminated government-operated facilities demands close attention (Forrest, 2003b). This is particularly important when neonate mortality is fully concurrent with high CWD prevalence, as is found in both the Colorado Division of Wildlife (CDOW) Ft Collins Foothills Research Facility (FWRF) and Colorado State University's Ft. Collins, Wildlife Disease Research Center (WDRC). Combined and coincident neonate mortality and CWD concurrence suggests that additional unrecognized disease transmission factors are at work (Neil, 1977, Miller, 1998, Williams & Young, 1980, 1982) Interestingly, similar high neonatal mortality incidents occurred at: 1) the old US Bureau of Sport Fisheries and Wildlife, Denver Wildlife Research Center (DWRC) (Robinette, 1973); 2) the US Forest Service's Frazer Experimental Forest deer pens (Neil, 1977); 3) the CDOW's Little Hills deer facility (Meeker, CO); 4) the CDOW's Junction Butte deer facility (Kremmling, CO) and; lastly at 5) the Author's domestic elk Ranch A (Forrest, 2003b). Significantly, all five facilities have either had a diagnosed CWD positive animal in residence, or were potentially exposed to CWD animals within their confines for extended periods of time. Sporadic symptoms found in suffering neonates are intermittently repeated in adult animals, generally within the last year of life prior to succumbing to CWD. These symptoms include: conjunctivitis, mucosal ulcers or abscesses both of the oral cavity and the stomach, nasal discharge, lameness and pneumonia. As such the co-incidence of high neonatal mortality and adult CWD, many times with corresponding subsequent symptoms cannot be ignored. Quite logically, a common agent must be involved, or lesser perhaps, there is a close commensal relationship between differing, yet symbiotic pathologic agents. Seemingly, two hazy or indistinct versions or stains of CWD may be at play within the wild and domestic cervids. The first strain is deemed CWDe (environmental). CWDe is manifested by an agent residual in the environment, but most likely ensconced in an insect host. The second, less distinct and of limited presence, shall be deemed CWDr (respiratory). CWDr is apparently a more virulent version of CWDe capable of severely impacting neonatal cervidea, in turn producing a more pronounced and sustained CWD epidemic. One must be aware, however, that although distinct and possible variant or stain characteristics are presented, the evidence of actual strains is not well developed. The manifested physical characteristics may not be so readily defined as biological strains, but simply as transmission strains representing an expression of the mechanism by which transference occurs. Certainly in the case of Ranch A, a presumed terminal CWDe heifer gave rise to a postulated CWDr calf, which then infected several dozen other elk calves with the more virulent, fatal neonatal version, CWDr. The CWDr agent seems to have passed via close animal-to-animal contact. One must recognize that upon the premises of a CWDr infection, the agent is likely to be returned to the environment via feces or urine so as to become available as CWDe. Perhaps eventually taking up residence in an extended secondary insect vector prior to introducing infection to a new cervid host at some future date. Table I demonstrates the characteristics of each suspected agent strain. TABLE I, Postulated CWD Strains
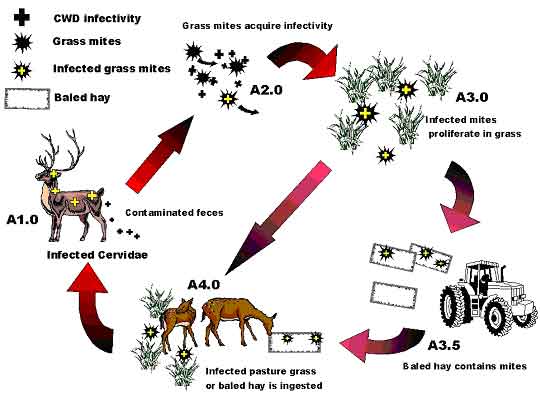
*Excluding fawn mortality in the first 24 hours after birth Infectivity Certainly, few will doubt the experimental transmissibility of the CWD agent via the alimentary system. This is the fundamental precept behind the CWDe infection mechanism. Basically, the agent is introduced orally into a proposed host via ingestion of a disease-bearing substance. Sigurdson (1999) documented the early presence (42 days) of infected prion-bearing alimentary lymphoid tissue upon oral ingestion of CWD-infected brain homogenate. However, the positive infection rate is suspected to be dosage dependant. The more ingested, the greater likelihood of sustainable infection. This suggests that the alimentary system itself has a finite, natural resistance to agent survival, much like that for many other biological pathogens. This recognized, minimum dose, oral infection process is the basis for most experimental CWD transmission research, not only in Cervids, but in Bovine and Ovid species as well. As such, the introduced agent seemingly enters the oral, esophageal, stomach, and/or the intestinal mucosa lining, overwhelming natural defenses and ultimately finding its way to the lymphatic system where chronic infection begins. Once its presence is established in the mucosal lining, the usefulness of direct alimentary system antibiotic treatment seemingly becomes erratic or useless. The presence of abundant infectivity in the gut lymphatic system at an early stage of TSE disease suggests the errant possibility of agent fecal shedding early in the disease progression (Radebold, 2001). Infectivity via the CWDr route is much less well known, let alone fully documented. The Ranch A experience suggests that the agent is easily passed between neonatal cervids, and has fully incubated within 7 days to instigate on-going contagious symptoms, which commonly result in short-term neonatal mortality. Unfortunately, no specific long-term studies have documented a progression from the limited calf survivability to full CWD in adult animals, however the circumstantial evidence is compelling. The most obvious suggested source of CWDr virulency and widespread infectivity is via the ever-present clear to yellow nasal discharge observed a few days prior to neo-natal collapse and short-term death. However, one cannot rule out infectivity obtained by exposure to diarrhea, solid feces or contaminated saliva via cross mothering, or from fawn dirt-eating, as is common in cervids kept in close confinement. Interestingly, as a side note, Fleetwood & Furley (1990) document TSE-infected elands (a member of the antelope family) at an English zoo. Initial symptoms included high stepping, muscular tremors and loss of weight, followed by circling, head pressing, a clear nasal discharge, hyper-salivation and recumbency preceding death after only 8 days of clinical signs. Initial symptoms are suggestive of encephalitis, while the nasal discharge mimics a Ranch A symptom. Dosage In the case of CWDe, the indigestion of the agent directly, or as it is found in a secondary insect vector, provides the same mechanism of infection as defined by experimental procedures. An infection is highly dependant upon dosage, due primarily to a seemingly strong immuno-resistance conveyed by the corrosive alimentary system itself and of course, the immediate backup lymphatic defenses enveloping the alimentary system. The erratic nature of the agent in the environment, particularly if in individual, discrete insects generally precludes a sustained heavy dose, but the agent is made available in erratic bursts of infectivity, which cumulatively, may or may not be of sufficient strength and abundance to overwhelm the host defense mechanisms. The outcome is then outwardly sporadic incident rates usually tied to young animals or those that have undergone unrelated stressful events, hence, have compromised immune defenses. With CWDr it would seem that in adults, the immune system is sufficiently capable of defeating the agent under most circumstances, wholly dependant upon the degree of standard immuno-resistance present within the animal. An immuno-suppressed animal would be at greater risk. This resistance may be directly affected by previous exposure to low-dose agent via CWDe or CWDr processes. Prior exposure at lesser dosage most likely creates a classical, but as yet, undiscovered antibody-based resistance. In a neonatal cervid, immuno-resistance to CWDr is expectedly quite low, or even lacking in ability to resist the pathogenic agent, hence, infection rates are high. Neo-natal mortality occurs rapidly, or a chronic condition ensues, eventually culminating in full-blown CWD many months later. The presence of a natural neonatal immunity conveyed either in utero or via colostrum is indicated by the elk calf survivability from non-clinical, but CWD-infected dams as documented by Miller (1998). However, elk herd disease statistics in South Dakota alternatively suggests specific maternal infection similar to that documented in sheep (Ritterhouse, G., PC, 2003). In respect to Miller (1998), the facilities at which the cervids were kept is arguably the most potentially CWD-polluted of any encountered, hence any animal exposed to that environment will likely commence antibody production immediately and perhaps can convey such immune response to it's offspring, despite eventually succumbing to the overwhelmingly present disease agent. Certainly in the suspected Ranch A experience, a clinical CWD dam (Cow #1) may convey disease directly to its unborn offspring (Calf #1c)(Forrest, 2003b). Growth Runting syndrome is apparent in both CWDe and CWDr. After the FWRF decontamination effort in 1985, the first three new CWD cases occurred among the three smallest 1986 herd cohorts (Miller, 1998). This parallels the findings at Ranch A where the initial CWD-infected 1999 elk heifer (Cow #1) was the smallest of 16 imported cohorts (Forrest, 2003b). Cow #1, incidentally, give birth to calf #1c, the suspected "Typhoid Mary" who later infected the remaining neonates with a virulent, fatal disease. Upon Ranch A depopulation, a second, smallish, mousey, Ranch-A-raised 1999 heifer was similarly diagnosed in a pre- or sub-clinical CWD state (Cow #3). Interestingly, Cow #3 bred to Bull #2 (also in a subclinical CWD stage) produced a very healthy calf, which did not succumb in any fashion (no symptoms expressed) to the calf pathogen, which killed 60% of its pre- and post-contemporaneous birthing cohorts. Of the surviving Ranch A 2001 calf crop, several had expressed the typical nasal discharge symptoms, but had eventually repressed the symptoms of the neonatal mortality pathogen. However, those survivors visually did not gain weight as fast, nor react as spryly as those who had not expressed any symptoms. Unfortunately, these survivors were only observed for a few weeks before they were summarily terminated by USDA order, that despite being preserved from earlier depopulation efforts by the urgings of the Author and Colorado State Vet's office. Critically, runting is an adjunct symptom visible prior to full CWD clinical manifestation. However, not all animals express runting, so it seems that the level of infectivity and manifestation is, at least at early stages, specifically animal dependant, some expressing, some not. Certainly the presence of runting suggests early infection and a disruption of normal growth processes, perhaps due to a chronic low-level infection diverting required energy and nutrients to internally motivated, prophylactic immune-related bacterial suppression processes. Significantly, runting has been documented in young animal chronic Spiroplasma infections. Select strains of Spiroplasma are able to survive up to nine months in intracerebrally inoculated mice and were associated with significant runting syndrome with an increase incidence of mortality and neurological symptoms generally without the appearance of antibody mimicking and immunological tolerance (Chastel, 1991). Bastian (1997) demonstrated that subcutaneous inoculation of rats with the Spiroplasma mirum GT-48 strain produced hair loss and a diminishment in body weight (runting), yet produced both unilateral and bilateral cataracts. Such was the case at Ranch A (Forrest, 2003b). POSTULATED CWD TRANSMISSION ROUTES Environmental Transmission From the examination of case data, Forrest (2003b) determined an outwardly appearing random and sporadic pattern of CWD occurrence within both within domestic facilities and amongst the wild cervidea population. Unfortunately to date, predictable and unambiguous disease transmission has only been documented under strictly controlled laboratory conditions. The most common and documented experimental transmission of TSE disease (other than by cerebral injection) is via ingestion of contaminated tissue believed to contain the causative pathogen. An early and substantial presence of the abnormal prion protein, a product of infection, within the alimentary system lymph nodes suggests that ingestion is potentially a significant factor in disease transmission. One logically must explore the role of a consumed agent in the natural transmission of CWD. As demonstrated by widespread disease dissemination in the Colorado endemic area, numerous occasions have arisen whereby the disease pathogen could depart an infected animal and potentially become residual in the environment. One can easily envision the feces or urine of a cervid containing the suspected CWD Spiroplasma bacteria, or other potential agent, deposited in a grassy field or pasture. Excretion debris would then logically represent a potentially infective source for grass, grain or forage mites, rummaging around in most pastoral environments. A mite passing across the debris, or feeding upon excreted plant material would contract or ingest the suspected bacteria. The microorganisms then became resident in the mites body, multiplying and re-multiplying, eventually being deposited with the eggs of the mites, leading to subsequent generations of mite infection. Each mite generation travels a few feet out from its original source, or perhaps is blown several tens of feet away by high winds, both ways progressively building the sphere of potential CWD environmental contamination. Infected mite concentrations would progressively increase in proportion against non-contaminated mites until such time as infective mites are present in ample quantities. At this point, an unfortunate new cervidea host consumes a sufficient quantity of such mites, leading to the release of infectious bacteria within the digestive system of the new host animal. CWD is the gradual result of the bacteria's systemic onslaught. Research studies have demonstrated high levels of infection found in the mesenteric and digestive lymph node tissue in both young and old animals. Logically, this leads to the conclusion that an oral route was most likely the major route of infection, particularly in the older animals. One must, however, also recognize that in a limited number of cases, the olfactory lymph nodes have also been intensely affected and may suggest an alternate infection scenario via nose-to-nose transmission. As demonstrated by the evidence presented in Forrest (2003b), domestic cervid herds have "out-of-the-blue" CWD infections wholly compatible with a random, feed-introduced pathogen source. Similarly wild and domestic cervids have demonstrated random infection when placed upon or adjacent to pastures previously containing CWD-infected animals. In most cases, such animals have advanced CWD symptoms within alimentary tract lymph nodes, suggesting that the most likely route of infection was via oral ingestion and assimilation through the alimentary mucosal lining. Interestingly, the occasionally recurrent, but normally infrequent presence of oral, reticular and abomasal ulcers in CWD affected animals, particularly neonates, suggests that the CWD pathogen may have an adverse affect upon these organ linings, membranes generally regarded as the first lines of defense against bacterial invaders. Equally important, the strong presence of CWD infectivity in the alimentary tract lymph nodes also infers the possibility that the agent may be readily excreted from the alimentary tract throughout the progression of the disease. This, in fact, may be the route of the greatest flushing of infective quantities of the pathogen due solely to the close proximity of a high infection area (the lymph nodes) to a potential route of excretion and hence, shedding of the agent. While observational studies suggest that agent excretion is very limited in the early stage of disease (Miller, 1998), its presence in the alimentary tract early in the disease process betrays that shedding of the agent probably does occur at all stages of disease, but is perhaps not of sufficient quantities to be deemed appropriately infective. However, once ensconced in mites or other insect, infectivity remains residual and reproducing in that particular vector until consumed by a susceptible host, next week, next month, or up to many years later, or most notably, is potentially harvested during commercial hay operations. Reasonably, the ingestion of a potential CWD pathogen can be easily accomplished in domestic herds through the use of commercially produced, mite-infested hay. Such hay being derived from fields frequented by diseased wild cervidea shedding infected feces, urine or saliva, or even by dead, rotting carcass debris. Transitory astigmatic mites then conceivably acquire the agent via contact or consumption remaining resident in the hay until consumed by an unfortunate cervidea. Such stored forage, particularly if left with sufficient moisture to propagate mold, may in fact create a microenvironment, quite conducive to the procreation of mites, enhancing infectivity by rapidly propagating infective mites and hence increasing dosage available for the unfortunate cervid. See Figure 1, which diagrammatically portrays the potential CWDe environmental mite reservoir cycle.
Figure 1: Proposed CWDe Environmental Reservoir Cycle.
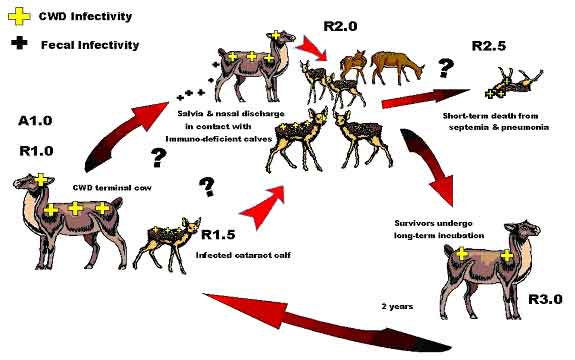
One must recognize that an environmental reservoir based upon a particular insect population is subject to some variation within a particular climatic region, as well as being affected by substantial variation between widely differing climates and eco-systems. Obviously the attendant insect assemblages differ greatly between dry deserts and wet, temperate rain forests. As such, one can expect the exact contaminated insect species or group of species in any one region may vary from those in another. Even the type of insect might vary from mites on the great grassy plains to mosquitoes on the frozen tundra. Much more work will need to be done to classify potential insect vectors. For instance, here in Colorado, just the elevation and climate peculiarities between the eastern and western slopes of the Colorado Rocky Mountains present unique differences. The eastern slope where CWD is widely recognized varies from the high plains, tall grass savanna at elevations of 4000 feet to about 6500 feet, up through into the coniferous forest zone at 6500 feet to over 9500 feet, and on, up into the Continental Divide alpine zone well over 10,000 feet. While insect assemblages can be expected to be gradational, some basic similarities, but perhaps some major differences are to be anticipated between those at 5000 feet and those at 10,000 feet. Quite simply put, the potential insect vector at 10,000 feet, if present at all, may be the same, partially similar, or totally different than the possible vectors present at sea level. Importantly, as could be expected if insect-vector ensconced, the rate of CWD prevalence will parallel climate as a function of elevation differences. Elevation certainly has a strong influence upon insect diversity and abundance, so why not upon the CWD infection rate of wild cervids as well? The vast majority of CWD mule deer cases are found in the tall grass savanna (prairie grasslands) of Wyoming and Colorado, and the associated man-made cultural modifications thereof. This high rate area predominantly occurs between the elevations of 5000 feet to perhaps 6500 feet, aligning north-south along eastern slope of the Colorado-Wyoming Front Range Mountains. This also corresponds to the rich irrigated farmlands of the South Platte River in the Front Range area or the North Platte River drainage in southern Wyoming. The higher CWD prevalance area in Wyoming also approximately mimics this Colorado north south trend paralleling the low mountains of the Laramie Range, but at generally less than the 7000-foot elevation. The tall-grass savanna grasslands are a particular ecotype obviously containing abundant tall range grass and other scrub-type vegetation. One would expect a similarity of insect species paralleling the general geomorphic and climatic trends. Further to the west, coniferous forests possessing pines, spruces, firs, etc. show much lesser quantities of grasses except in the areas of open meadows, a la, Estes Park, which by definition, is an open park containing abundant meadow grasses and extraordinarily high CWD prevalence. Elk cases are found predominately at the contact between the forests and the grasslands reflecting the elk's desire to have readily available forest cover. While certainly grasslands are found to the west side of the divide, the elevations are considerably different. The western slope's Middle Park area is above 7000 feet to over 9000 feet and although abundant grassy meadows are present, much of the surrounding area is all well above 9000 feet. As such, a tangibly different climate is found on the west side of the mountains from the east side. That climate affects vegetation, which in turn affects insect diversity and abundance. Still farther to the west, a drier climate means a lesser abundance of vegetation, which in turn means lesser insect inhabitants. Lesser insects yield a lesser rate of potential infection due to fewer animals grazing per acre. So under the solely ideal CWDe mode of transmission, a drier climate would likely have a lesser rate of infection. Since elevation-related climatic variations will influence the distribution of the insect vector, and hence the availability of disease, then any condition, which would enhance the proliferation of the insect vector would correspondingly also enhance the prevalence of disease. Perhaps the warm and mild winters in Colorado and Wyoming over the last decade have led to less direct insect winter kill and hence, to the development of more abundant affected mite or insect populations, basically a fore runner or predilection of disease prevalence amongst the resident cervidea. Respiratory Transmission Evidence of a possible respiratory route of Spiroplasma infection can be gleaned from the examination of the role of the closely related, more studied mycoplasmas defined in lesser-known diseases. While mycoplasma and Spiroplasma are cousins, they are not identical in characteristics. Spiroplasma with its helical shape is motile while mycoplasmas are not. This renders mycoplasmas less mobile and more dependant upon host proclivities for transport and infection. In Germany, Kern (1998) documented the only known case of a human Spiroplasma infection, describing cataract development and a bout of pneumonia of unknown, but a suspected spiroplasmal cause. Certainly everyone has heard of "walking" pneumonia. A chronic respiratory condition (atypical pneumonia) with prolonged symptoms and a resistance to treatment, which can be debilitating but is not necessarily fatal. This is the most common form of pneumonia among humans age 5 to 35, and is believed caused by Mycoplasma pneumoniae. Similar contagions affect swine (Mycoplasma hyopneumoniae or M. hyorhinis), goats (M. mycoides capri), cattle (M. mycoides mycoides), and sheep (M. ovipneumoniae). The sheep variety is particularly telling, as it can be clinically expressed by nasal discharge, breathing irregularities and mild fever, followed by "runting" in young victims. All can be passed via aerosols from animal to animal. A particularly pertinent M. mycoides mycoides (large colony) infection in New Zealand dairy goatherds has been well documented (Jackson, 2001). Three to 20 day-old goat kids expressed polyarthritis, fever and septicemia. Exposed bovine dairy calves developed polyarthritis, septicemia and conjunctivitis. Some calves and kids developed fatal pneumonia. The source of infection was determined to be infected milk or colostrum. Earlier M. mmlc transmission studies found that upon experimental oral dosage, 37 of 51 (72.5%) one-week-old kids fatally succumbed, as did 8 of 14 (57%) in-contact, but non-dosed control kids (DaMassa, 1986). No doubt that transmission could occur via close, nose-to-nose contact, but it is difficult to gauge the importance of the respiratory route of infection since it cannot be easily distinguished from the oral or oral-pharyngeal route (Jackson, 2001). Jackson further noted that mycoplasma infection should be suspected in goat kids when presented with (a) sudden death, (b) central nervous system spasm with opisthotonus (body bowed down or forward, concave backbone) and death within 24 to 72 hours, (c) fever and septicemia, as well as polyarthritis, lameness, anorexia or pneumonia. Affirmatively, some of the suspected mycoplasma infection criteria, particularly septicemia and opisthotonus, were found during Ranch A's 2001 elk neonatal mortality incident (Forrest, 2003b). Essentially, the respiratory route of infection is seemingly one of closely confined animals, whereby direct animal-to-animal contact is available on a day-to-day extended basis. This situation is not common under purely wild conditions. Seemingly, the ability to have direct contact, or be within close proximity of mucosal aerosol dispersion is critical for disease propagation via the respiratory route. As such, it is not believed to be a major dispersion route in the wild or even among domestic cervid herds, but is applicable only to selected situations where the individual animal symptoms and group herd circumstances are conducive to transmission. Obviously, the closer stocking rates in domestic facilities, or in wild captive research facilities can precipitate a respiratory infraction, however, concentration of wild animals around isolated watering holes or due to artificial feeding situations could conceivably exacerbate the prospective problem as well. Neonates are apparently much more susceptible to infection via the respiratory system. Most likely, this is due to their immature immune system, lacking disease antibodies, and their small size, making lethal dosage allotment more certain, together with bacterial colonization more probable. The extended survivability of a few elk calves retained upon bovine colostrum at Ranch A suggests that antibodies to the CWD agent or a close cousin may be present in bovine colostrum, while not present in normal cervid colostrum. An adult version of the respiratory infection route seems much less common, but cannot be eliminated altogether. Perhaps the normal adult immune function is usually sufficient to repulse the pathogen, or perhaps the dosage gained by brief contact is insufficient to take hold. Whatever the reason, adult respiratory infection may be limited to immuno-suppressed individuals sustaining contemporaneous parasite infections or other debilitating problems. The difficulties in segregating adult respiratory infections whether they be from oral or ingested infections, makes full differentiation of the route of transmission virtually impossible, as was found by Jackson (2001) in reference to the New Zealand mycoplasma infections. Certainly, amongst Ranch A elk calves, a birthing group confined under high stocking rates, the most obvious route of respiratory infection was via direct contact with the ever present, clear to yellowish, potentially infective, nasal discharge so prevalent among the affected calves. A nasal discharge is potentially a lesser-recognized cervid symptom of TSE or CWD infections, particularly among neonates, but has also been documented in the CDOW's FWRC pens in late-stage, clinical CWD animals. As noted earlier, Fleetwood & Furley (1990) document TSE-infected elands having a clear nasal discharge prior to a TSE-induced death. Such discharge contact may be via direct nose-to-nose contact between spry, inquisitive neonates, or more subtly by indirect contact via shared udder contact allowed by new mothers, or perhaps even shared grooming by cohorts or foster-care mothers. Lastly, of course, transitory aerosol contact with dispersed, but still virulent, mucosal discharge is always possible with coughing or sneezing animals. While a nasal discharge might be generally ascribed to concurrent fibropurulent broncho-pneumonia or other similar malady, one must recognize that the pneumonia itself may actually be an early, mid- or late-stage symptom of the CWD pathogen, derived from a direct bronchial Spiroplasma infection, or perhaps from a mycoplasma commensal agent. Seemingly though, the nasal discharge found in Ranch A calves was not pneumonia-induced. Multiple necropsies failed to find consistent pneumonia symptoms. The calves displayed simply rhinitis without any other obviously related conditions, more akin to say a common cold, but with much more severe an outcome after several days. Figure 2, diagrammatically portrays the potential CWDr respiratory infection cycle
Figure 2: Postulated "Ranch A" Lethal Respiratory CWDr Infection Cycle
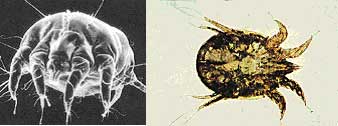
Effectively the respiratory infection route displayed at Ranch A, commenced with the birth of calf #1c, born out of the womb of a clinical CWD mother, complete with rhinitis and a blue cataract. Close contact with other curious calves, born contemporaneously or thereafter, proved fatal for many of them but not all. Much like the calf and fawn mortalities at the CDOW's FWRC and CSU's WDRC facilities, at which the surviving animals eventually succumbed to adult CWD, the CWD pathogen was emplaced within their bodies as neonates, or soon acquired it from cohorts or feed within a short time thereafter. Ectoparasite Mite Transmission Astigmatic grain and mold mites have been implicated as possible vectors in the transmission of TSE disease (Wisniewski, 1996, Rubenstein, 1998 and Carp 2000). Chastel (1987, 1991, 1992) demonstrated that Spiroplasma, a motile mycoplasma derived from the Class Mollicutes, is relatively abundant, albeit generally undetected, in the insect world. Elizan (1972) and Bastian demonstrated that Spiroplasma could cause symptoms compatible with TSE disease. Forrest (2002) determined that Spiroplasma retains the distinction of harmonizing with the most known TSE agent characteristics when present in vertebrate animals. Forrest (2003a) elaborated upon the particular role of astigmatic mites and how Spiroplasma bacteria could be the TSE causal agent within the mite vector, suggesting that bacteria would be capable of reproducing within the mite body, hence becoming capable of sustaining long-term environmental contamination. So, what other devious role might astigmatid mites play in the CWD drama unfolding across the North American continent? Astigmatic or Astigmatid refers to the lack of a "stigmatae", the tubular breathing apparatus used by the vast majority of Acari or "mites". Unfortunately, mite phylogenic classifications are extremely complex and sometimes contradictory. Suffice to say that of the some 5000 genera and 50,000 individual mite species that have been definitively identified around the world, Astigmata contains some 1054 genera and only 1131 individual species (Hallan, J., 2001, Web Pub: Synopsis of the Acari of the World). A few of those specific species (i.e., Lepidoglyphus destructor, Acarus farris, and Tyrophagus longior) were implicated in the potential transmission of TSE disease (Wisniewski, 1996, Rubenstein, 1998, Carp, 2000 and Forrest, 2003a). In addition to the prolific grain and mold mites, several additional common astigmatid mite genera are regarded as ectoparasites, minute parasitic creatures living on the outside of mammalian hosts. This includes such well-known parasites as psoroptes, sarcoptes and chorioptes mites collectively known as the "scabie" mites. Scabie mite infections of mammals are common. Under Federal law, scabie infections must be reported to the appropriate state control agency, and are subject to quarantine. At least 15 species of psoroptic mites and some 24 sarcoptic species have been identified (Hallan, J., 2001, Web Pub: Synopsis of the Described Mite Families Of The World). While the USDA claimed to have eliminated Psoroptic scabies in sheep by 1973 (USDA 1999 Agricultural Fact Book), Psoroptes continuing presence in wildlife and upon livestock has been frequently documented. Of these parasitic mites, the sheep psoroptic mange mite Psoroptes ovis, and its close associate, Psoroptes cunculi, both non-burrowing skin mites are of particular interest. P. ovis is believed to cause body mange (scabies), while P. cuniculi preferably infests ear canals causing canker. Interestingly, over many years, psoroptic mite infections were repeatedly noted at both the Colorado Division of Wildlife's Foothill Wildlife Research Center (FWRC) and at Colorado State University's Wildlife Disease Research Center (WDRC) at Ft. Collins, Colorado (CDOW deer records, and Neil, P., PC 2/2003). Roberts (1970) first documented Psoroptes cuniculi in mule deer at the USBSFW's Denver Wildlife Research Center, Lakewood Co. (DWRC) which had exchanged animals with the FWRC and WDRC. Pillmore, a co-worker of Roberts noted that domestic sheep, wild Rocky Mountain Bighorn sheep, hybrid domestic-Bighorn sheep, wild-caught mule deer and even rabbits were all in close residency at the DWRC, and may have created mite population cross-infections (Pillmore, R.E., PC, 2003). The FWRC and WDRC also had Bighorn and domestic sheep, mule deer, white-tailed deer, elk and wild rabbits. Strickland (1970) noted that white-tailed deer also can carry Psoroptic spp mites, as could Mexican Bighorn sheep (Lange, 1980). P. cervinus was noted as an ear mite of bighorn sheep and wapiti (Sweatman, 1958), but may be a variant of P. cuniculi. Conservatively, mites in the Genus Psoroptes should not be considered host-specific. They readily transfer between hosts (Sweatman, 1958). Wright et al. (1983) was successful in cross-mating bovine-derived P. ovis with rabbit-derived P. cuniculi. The reciprocal crosses were infective to both cattle and rabbits, and suggested that P. ovis and P. cuniculi were not reproductively isolated, but distinct races of the same mite, differentiated solely by host preferences. Similarly, Boyce (1990) noted that differences were not detected between mites collected from sympatric populations of infested mule deer and bighorn sheep, suggesting that these mites are not host specific, but represent a single interbreeding population. Differences also were not detected among mites collected from the ears and body of bighorn sheep and rabbits, demonstrating that the location of mites on a given host should not be used as a primary criterion in species identification. In fact, Boyce and Brown (1991) using immuno-blotting techniques with defined antigen and antisera demonstrated extensive and nearly complete antigen cross reactivity between various Psoroptes spp. isolated from bighorn sheep, mule deer, cattle and rabbits. Bates (1999) later suggested that Psoroptes infesting sheep (i.e. P. ovis and P. cuniculi) are not reproductively or ecologically isolated, but are phenotypic variants of the same species. Moving on, what role might psoroptic mites play? As previously discussed, the Mycoplasma mycoides mycoides (large colony) infections, as documented in New Zealand dairy goat kids, is a rare glimpse into the nature of less motile mycoplasmal infections. Both New Zealand and Australian M. mm case studies present compelling evidence of unique mycoplasmal transfer. Importantly, (Cottew &Yeats, 1981) detected non-motile Mycoplasmas in the external ear canal of goats by swabbing and culture. Up to 10(8) colony forming units were recovered from single swabs. Pathogenic species of M. agalactiae, M. capricolum, M. mycoides capri, M. mycoides subsp. lc (mycoides of the large colony (LC) type), M. putrefaciens and three un-typed mycoplasmas were present. Significantly, the same mycoplasmas were found in large numbers of resident mites, Psoroptes cuniculi and Raillietia caprae, present in the external ear canal. Cottew noted that the role of mycoplasmas in the external ear canal as a source of infection and disease, and of the mites in the spread of infection requires further elucidation. Interestingly, Hazell (1985) further documents the presence of M. mm in the ear canal of goats sustaining an outbreak of systemic mycoplasmosis. The adult female Psoroptes spp. is just visible to the naked eye, approximately 750 µm in length. When colonizing the ear canals of any hosts or upon the bodies of sheep, these mites are pearly white and globular in appearance, see Photo 1 a & b. However, they may appear black to dark red when in long term residence upon the bodies or exterior ear due to the ingestion of red blood cells (Wright and Deloach, 1980, 1981). Dark brown "coffee grinds" in the ear canal can suggest infestation.
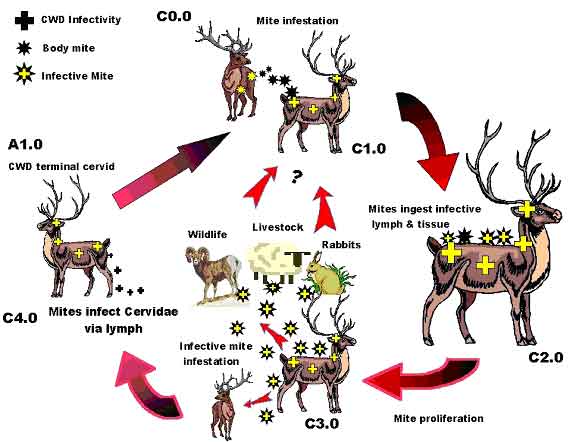
Photo 1 a & b: Psoroptes ovis, aka, P. cuniculi Psoroptic mites are fully parasitic, needing a suitable host for survival. Mite mouthparts are elongated and adapted for piercing skin to suck lymph fluid. All stages of their life cycle occur around the edges of their host's oozing lesions. Regeneration time is from 1.5 to 3 weeks, but Psoroptes can survive off the host for 2 to 3 weeks. They prefer areas of thick hair or in the ears, but untreated infections will spread over the entire body surface. Nibbling by these lymph-sucking mites cause inflammatory lesions that ooze fluid, which, in turn dry to form crusts (scabs, hence "scabies"). The bare scabby areas produced are unsuitable for the mites, which migrate to surrounding healthy skin, potentially spreading tissue damage over the entire body. Intense infections can cause emaciation and death. (http://nz.merial.com/horse_owners/disease/psoro.html) Mapstone (1982) suggested that the unusual nature of Psoroptes mouthparts might facilitate a crucial, two-way flow of liquid, where saliva flows down a central salivary canal and spills out onto the host's skin, where external serrated leg parts meet and abrade the skin. Liquid food may move along tubular grooves to be sucked up a food canal formed by the mouthpart walls. Notably, if Psoroptes can distribute saliva, Psoroptes can transfer potentially infective bacteria in its saliva into the host's open wounds, hence transferring a bacterial infection to a new host. Conversely, and just as important, the lymph-sucking nature of Psoroptes makes it an ideal vector for the inter-animal spread of blood-borne disease. Recalling the TSE infectivity data of Forrest (2002), lymph nodes are important sites of initial TSE discrimination through the recognition of abnormal prion protein. Tamai (1992) noted that leukocytes from a TSE-infected human mother were infective, as was the mother's colostrum, similar to M. mmlc dairy goat infections noted above. Klein (1998) additionally postulated that B-leukocytes might assist in the transport of scrapie infectivity. While whole blood was found to be infective, Holada (2002) found a larger proportion (98.4%) of the total infectivity was recovered from the mononuclear leukocyte fraction. So fundamentally, Psoroptes derives its sustenance from a highly infective TSE-agent bearing substance. This same substance, the white blood cell rich, lymph, is suspected to be the vehicle used by Spiroplasma to confound the host immune system and gain access to the central nervous system eventually resulting in TSE disease (Forrest, 2002). Seemingly, under close animal confinement conditions, the presence of "ear mites" and Chronic Wasting Disease may well go hand-in-hand. Figure 3 diagrammatically portrays the potential ectoparasite CWD infection cycle.
Figure 3: Postulated Cervidae Ectoparasite Mite Cycle.
Conclusively, a Psoroptes mite infection associated with the TSE infection process must be closely investigated. In our limited knowledge, certainly Psoroptes cuniculi can be regarded as an unknown, but potentially serious prospective transmission vector, one well worthy of a close and critical examination. If Psoroptes can play a role in CWD, can a deliciously inexorable link between scrapie and scabies be far behind? A continuing paper: Chronic Wasting Disease - Part IId Observations on TSE Transmissibility -"CWD Created?" will attempt to place all the previous information from prior papers into one grand hypothetical notion as to how and why CWD may have come into being, and where it might now be present. About the Author's CWD Foundation: In the summer of '01, the industry-mandated CWD surveillance program found a single CWD positive animal in the author's domestic elk herd. By February 2002 the entire +300-head herd was depopulated. The cumulative CWD knowledge amongst the entire scientific community was perceived to be minimal at best. For some 35 years the pathogen was virtually ignored by the research community until the ravages of Britain's Mad Cow Disease and the recognition of human-variant Creutzfeldt-Jakob Disease demanded closer attention. This compendium of papers, co-sponsored by The CWD Foundation is in clear recognition of a dire need for a better understanding of CWD epidemiology. Our hope is that creative discourse will help decipher the required solutions. We welcome financial support from concerned parties. Contact: Rich Forrest or Jan Elsworth, The CWD Foundation, Box 55, South Fork, CO 81154, telephone: 719-657-0942, email: Research@stopcwd.org. REFERENCES Bastian, F.O., et al, 1987, Ann Inst. Pasteur/Microbiol. V138, pp 651-655 |